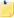PF Night Splint Compared to plantar based splints, patients prefer dorsal models because they are less bulky, cooler, lower profile and easier to walk in at night. PF Night Splint includes all of these features and more. Neoprene calf and toe straps along with gel padding on the top of the foot help reduce pressure points and the plastic-free bottom offers safer nighttime ambulation. Even added more room in the forefoot to accommodate wider feet and new flexible shell allows easier nighttime ambulation without compromising the ability to hold the foot at 90° during rest. Finally, a plantar fasciitis splint patients will wear all night! Neoprene calf. Padded foam liner. Fits right or left. Not made with natural rubber latex.
PF Night Splint (Derotation)
Applications
Prevention and correction of foot drop.
Night splint for early healing.
Ambulatory, can be used as a day splint.
Perfect post-operative immobilisation and derotation.
Peroneal / Peritibial nerve or muscle damage.
Ankle or Plantar flexion contracture and functional alignment.
Can be used to protect the diabetic/ injured ankle & foot.
PF Night Splint (Derotation) Features
Removable de-rotation bar.
Moulded foot casing, aesthetically pleasing and durable.
Effective Liner, improved comfort.
Highly functional Design, customized degree of dorsiflexion.
Double strapping mechanism, better grip.
Tynor R O M Knee Brace
Prefabricated, custom adjusted R.O.M. Knee Brace controls and restrict flexion/extension of the knee joint through a multi-point joint fixation system to allow a range of motion or immobilization to the knee. Universal design allows it to be used on quite a big range of population and for a multiple orthopedic problems associated with knee joint.
Immobilization at any angle
Controlled motion between any two angles
Quick and easy application and removal.
Tynor R O M Knee Brace Features
R.O.M. Dial
0 to 120 degree flexion.
0 to 60 degree hyper extention
Locking at any
Around 50% of ageing, obese men presenting to the diabetes clinic have lowered testosterone levels relative to reference ranges
based on healthy young men
The absence of high-level evidence in this
area is illustrated by the Endocrine Society testosterone therapy in men with androgen deficiency clinical practice guidelines
(Bhasin et al. 2010), which are appropriate for, but not specific to men with metabolic disorders. All 32 recommendations made in these guidelines
are based on either very low or low quality evidence.
A key concept relates to making a distinction between replacement and pharmacological testosterone therapy
The presence of symptoms was more closely linked to increasing age than to testosterone levels
Findings similar to type 2 diabetes were reported for men with the metabolic syndrome, which were associated with reductions
in total testosterone of −2.2 nmol/l (95% CI −2.41 to 1.94) and in free testosterone
low testosterone is more predictive of the metabolic syndrome in lean men
Cross-sectional studies uniformly show that 30–50% of men with
type 2 diabetes have lowered circulating testosterone levels, relative to references based on healthy young men
In a recent cross-sectional study of 240 middle-aged men (mean age 54 years) with either type 2 diabetes, type 1 diabetes
or without diabetes (Ng Tang Fui et al. 2013b), increasing BMI and age were dominant drivers of low total and free testosterone respectively.
both diabetes and the metabolic syndrome are associated with a modest reduction in testosterone, in magnitude comparable
with the effect of 10 years of ageing
In a cross-sectional study of 490 men with type 2 diabetes, there was a strong independent association of low testosterone
with anaemia
In men, low testosterone is a marker of poor health, and may improve our ability to predict risk
probably the most important point made in this article
low testosterone identifies men with an adverse metabolic phenotype
Diabetic men with low testosterone are significantly more likely to be obese or insulin resistant
increased inflammation, evidenced by higher CRP levels
Bioavailable
but not free testosterone was independently predictive of mortality
It remains possible that low testosterone is a consequence of insulin resistance, or simply a biomarker,
co-existing because of in-common risk factors.
In prospective studies, reviewed in detail elsewhere (Grossmann et al. 2010) the inverse association of low testosterone with metabolic syndrome or diabetes is less consistent for free testosterone
compared with total testosterone
In a study from the Framingham cohort, SHBG but not testosterone was prospectively and independently associated with incident
metabolic syndrome
low SHBG (Ding et al. 2009) but not testosterone (Haring et al. 2013) with an increased risk of future diabetes
In cross-sectional studies of men with (Grossmann et al. 2008) and without (Bonnet et al. 2013) diabetes, SHBG but not testosterone was inversely associated with worse glycaemic control
SHBG may have biological actions
beyond serving as a carrier protein for and regulator of circulating sex steroids
In men with diabetes, free testosterone, if measured by gold standard equilibrium dialysis (Dhindsa et al. 2004), is reduced
expensive, laborious process filled with variables
Low free testosterone remains inversely associated with insulin resistance, independent of SHBG (Grossmann et al. 2008). This suggests that the low testosterone–dysglycaemia association is not solely a consequence of low SHBG.
Experimental evidence reviewed below suggests that visceral adipose tissue is an important intermediate (rather than a
confounder) in the inverse association of testosterone with insulin resistance and metabolic disorders.
testosterone promotes the commitment of
pluripotent stem cells into the myogenic lineage and inhibits their differentiation into adipocytes
testosterone regulates the metabolic functions
of mature adipocytes (Xu et al. 1991, Marin et al. 1995) and myocytes (Pitteloud et al. 2005) in ways that reduce insulin resistance.
Pre-clinical evidence (reviewed in Rao et al. (2013)) suggests that at the cellular level, testosterone may improve glucose metabolism by modulating the expression of the glucose-transported
Glut4 and the insulin receptor, as well as by regulating key enzymes involved in glycolysis.
More recently testosterone has
been shown to protect murine pancreatic β cells against glucotoxicity-induced apoptosis
Interestingly, a reciprocal feedback also appears to exist, given that not only chronic (Cameron et al. 1990, Allan 2013) but also, as shown more recently (Iranmanesh et al. 2012, Caronia et al. 2013), acute hyperglycaemia can lower testosterone levels.
There is
also evidence that testosterone regulates insulin sensitivity directly and acutely
In men with prostate cancer commencing androgen deprivation therapy, both total as well as, although not in all studies (Smith 2004), visceral fat mass increases (Hamilton et al. 2011) within 3 months
More prolonged (>12 months) androgen deprivation therapy has been associated with increased risk of diabetes in several
large observational registry studies
Testosterone has also been shown to reduce the concentration of pro-inflammatory cytokines in some, but not all studies, reviewed
recently in Kelly & Jones (2013). It is not know whether this effect is independent of testosterone-induced changes in body composition.
the observations discussed in this section suggest that it is the decrease in testosterone that causes insulin
resistance and diabetes. One important caveat remains: the strongest evidence that low testosterone is the cause rather than
consequence of insulin resistance comes from men with prostate cancer (Grossmann & Zajac 2011a) or biochemical castration, and from mice lacking the androgen receptor.
Several large prospective studies have shown that weight gain or development of type 2 diabetes is major drivers of the
age-related decline in testosterone levels
there is increasing evidence that healthy ageing by itself is generally not associated with marked reductions in
testosterone
Circulating testosterone, on an average 30%, is lower in obese compared with lean men
increased visceral fat is an important component in the association of low testosterone and insulin resistance
The vast majority of men with metabolic disorders have functional gonadal axis suppression with modest reductions in testosterone
levels
obesity is a dominant
risk factor
men with Klinefelter syndrome have an increased risk of metabolic disorders. Interestingly, greater body fat
mass is already present before puberty
Only 5% of men with type 2 diabetes have elevated LH levels
inhibition of the gonadal axis predominantly takes place in the hypothalamus,
especially with more severe obesity
Metabolic factors, such as leptin, insulin (via deficiency or resistance) and ghrelin
are believed to act at the ventromedial and arcuate nuclei of the hypothalamus to inhibit gonadotropin-releasing hormone (GNRH)
secretion from GNRH neurons situated in the preoptic area
kisspeptin has emerged as one of the
most potent secretagogues of GNRH release
hypothesis that obesity-mediated inhibition of kisspeptin signalling contributes to the suppression
of the HPT axis, infusion of a bioactive kisspeptin fragment has been recently shown to robustly increase LH pulsatility,
LH levels and circulating testosterone in hypotestosteronaemic men with type 2 diabetes
A smaller study with a similar experimental design found that acute testosterone withdrawal reduced insulin sensitivity
independent of body weight, whereas oestradiol withdrawal had no effects
suppression of the diabesity-associated HPT axis is functional, and may hence be
reversible
Obesity and dysglycaemia and associated comorbidities such as obstructive sleep apnoea (Hoyos et al. 2012b) are important contributors to the suppression of the HPT axis
weight gain and development
of diabetes accelerate the age-related decline in testosterone
Modifiable risk factors such as obesity and co-morbidities are more strongly associated with a decline in circulating testosterone
levels than age alone
55% of symptomatic androgen deficiency reverted to a normal testosterone or an asymptomatic state after 8-year follow-up,
suggesting that androgen deficiency is not a stable state
Weight loss can reactivate the hypothalamic–pituitary–testicular axis
Leptin treatment resolves hypogonadism in leptin-deficient men
The hypothalamic–pituitary–testicular axis remains responsive to treatment with aromatase inhibitors or selective oestrogen
receptor modulators in obese men
Kisspeptin treatment increases LH secretion, pulse frequency and circulating testosterone levels in hypotestosteronaemic men
with type 2 diabetes
change in BMI was associated with the change in testosterone (Corona et al. 2013a,b).
weight loss can lead to genuine reactivation
of the gonadal axis by reversal of obesity-associated hypothalamic suppression
There is pre-clinical and observational evidence that chronic hyperglycaemia can inhibit the HPT axis
in men who improved their glycaemic control over time, testosterone levels increased. By
contrast, in those men in whom glycaemic control worsened, testosterone decreased
testosterone levels should be measured after successful weight loss to identify men with an insufficient rise
in their testosterone levels. Such men may have HPT axis pathology unrelated to their obesity, which will require appropriate
evaluation and management.
Trends tell a lot more than individual values. IN the battle against obesity, trends tell a sad story. Obesity rates are increasing in our children. Up to 17.2% in children. The numbers in the severe category fared the worse.
Study finds salivary 8-OHdG is associated with increased severity of ED in men. The study found that phosphodiesterase 5 inhibitor therapy increased total and free Testosterone as well as decreased 8-OHdG.
Patients with ventricular tachycardia demonstrated significantly lower serum values of FT3 and FT3/FT4, and significantly higher values of rT3. Serum thyroid hormone levels can provide a quantitative index for evaluating the severity of chronic heart failure and predicting ventricular tachycardia.
The most striking abnormality detected in 75 sick euthyroid patients was a highly significant reduction in the mean total serum-triiodothyronine (T3) levels, with most patients having total T3 levels in the hypothyroid range. The severity of the illness correlated well with the reduction in total serum-T3 levels.
Acquired generalized lipodystrophy (AGL) is marked by severe insulin resistance and hypertriglyceridemia. Rarely, AGL and type 1 diabetes (T1D) coexist.
Long-term recombinant leptin therapy is effective in treating the insulin resistance of patients with the unusual combination of T1D and AGL.
Long-chain polyunsaturated fatty acids, including eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), are highly enriched in neuronal synaptosomal plasma membranes and vesicles
The predominant CNS polyunsaturated fatty acid is DHA
effective supplementation and/or increased ingestion of dietary sources rich in EPA and DHA, such as cold-water fish species and fish oil, may help improve a multitude of neuronal functions, including long-term potentiation and cognition.
multiple preclinical studies have suggested that DHA and/or EPA supplementation may have potential benefit through a multitude of diverse, but complementary mechanisms
pre-injury dietary supplementation with fish oil effectively reduces post-traumatic elevations in protein oxidation
The benefits of pre-traumatic DHA supplementation have not only been independently confirmed,[150] but DHA supplementation has been shown to significantly reduce the number of swollen, disconnected and injured axons when administered following traumatic brain injury.
DHA has provided neuroprotection in experimental models of both focal and diffuse traumatic brain injury
potential mechanisms of neuroprotection, in addition to DHA and EPA's well-established anti-oxidant and anti-inflammatory properties
Despite abundant laboratory evidence supporting its neuroprotective effects in experimental models, the role of dietary DHA and/or EPA supplementation in human neurological diseases remains uncertain
Several population-based, observational studies have suggested that increased dietary fish and/or omega-3 polyunsaturated fatty acid consumption may reduce risk for ischemic stroke in several populations
Randomized control trials have also demonstrated significant reductions in ischemic stroke recurrence,[217] relative risk for ischemic stroke,[2] and reduced incidence of both symptomatic vasospasm and mortality following subarachnoid hemorrhage
Clinical trials in Alzheimer's disease have also been largely ineffective
The clinical evidence thus far appears equivocal
curcumin has gained much attention from Western researchers for its potential therapeutic benefits in large part due to its potent anti-oxidant[128,194,236] and anti-inflammatory properties
Curcumin is highly lipophilic and crosses the blood-brain barrier enabling it to exert a multitude of different established neuroprotective effects
in the context of TBI, a series of preclinical studies have suggested that pre-traumatic and post-traumatic curcumin supplementation may bolster the brain's resilience to injury and serve as a valuable therapeutic option
Curcumin may confer significant neuroprotection because of its ability to act on multiple deleterious post-traumatic, molecular cascades
studies demonstrated that both pre- and post-traumatic curcumin administration resulted in a significant reduction of neuroinflammation via inhibition of the pro-inflammatory molecules interleukin 1β and nuclear factor kappa B (NFκB)
no human studies have been conducted with respect to the effects of curcumin administration on the treatment of TBI, subarachnoid or intracranial hemorrhage, epilepsy or stroke
studies have demonstrated that resveratrol treatment reduces brain edema and lesion volume, as well as improves neurobehavioral functional performance following TBI
green tea consumption or supplementation with its derivatives may bolster cognitive function acutely and may slow cognitive decline
At least one population based study, though, did demonstrate that increased green tea consumption was associated with a reduced risk for Parkinson's disease independent of total caffeine intake
a randomized, placebo-controlled trial demonstrated that administration of green tea extract and L-theanine, over 16 weeks of treatment, improved indices of memory and brain theta wave activity on electroencephalography, suggesting greater cognitive alertness
Other animal studies have also demonstrated that theanine, another important component of green tea extract, exerts a multitude of neuroprotective benefits in experimental models of ischemic stroke,[63,97] Alzheimer's disease,[109] and Parkinson's disease
Theanine, like EGCG, contains multiple mechanisms of neuroprotective action including protection from excitotoxic injury[97] and inhibition of inflammation
potent anti-oxidant EGCG which is capable of crossing the blood-nerve and blood-brain barrier,
Epigallocatechin-3-gallate also displays neuroprotective properties
More recent research has suggested that vitamin D supplementation and the prevention of vitamin D deficiency may serve valuable roles in the treatment of TBI and may represents an important and necessary neuroprotective adjuvant for post-TBI progesterone therapy
Progesterone is one of the few agents to demonstrate significant reductions in mortality following TBI in human patients in preliminary trials
in vitro and in vivo studies have suggested that vitamin D supplementation with progesterone administration may significantly enhance neuroprotection
Vitamin D deficiency may increase inflammatory damage and behavioral impairment following experimental injury and attenuate the protective effects of post-traumatic progesterone treatment.[37]
emerging evidence has suggested that daily intravenous administration of vitamin E following TBI significantly decreases mortality and improves patient outcomes
high dose vitamin C administration following injury stabilized or reduced peri-lesional edema and infarction in the majority of patients receiving post-injury treatment
it has been speculated that combined vitamin C and E therapy may potentiate CNS anti-oxidation and act synergistically with regards to neuroprotection
one prospective human study has found that combined intake of vitamin C and E displays significant treatment interaction and reduces the risk of stroke
Pycnogenol has demonstrated the ability to slow or reduce the pathological processes associated with Alzheimer's disease
Pcynogenol administration, in a clinical study of elderly patients, led to improved cognition and reductions in markers of lipid peroxidase
One other point of consideration is that in neurodegenerative disease states like Alzheimer's disease and Parkinson's disease, where there are high levels of reactive oxygen species generation, vitamin E can tend to become oxidized itself. For maximal effectiveness and to maintain its anti-oxidant capacity, vitamin E must be given in conjunction with other anti-oxidants like vitamin C or flavonoids
These various factors might account for the null effects of alpha-tocopherol supplementation in patients with MCI and Alzheimer's disease
preliminary results obtained in a pediatric population have suggested that post-traumatic oral creatine administration (0.4 g/kg) given within four hours of traumatic brain injury and then daily thereafter, may improve both acute and long-term outcomes
Acutely, post-traumatic creatine administration seemed to reduce duration of post-traumatic amnesia, length of time spent in the intensive care unit, and duration of intubation
At three and six months post-injury, subjects in the creatine treatment group demonstrated improvement on indices of self care, communication abilities, locomotion, sociability, personality or behavior and cognitive function when compared to untreated controls
patients in the creatine-treatment group were less likely to experience headaches, dizziness and fatigue over six months of follow-up
CNS creatine is derived from both its local biosynthesis from the essential amino acids methionine, glycine and arginine
Studies of patients with CNS creatine deficiency and/or murine models with genetic ablation of creatine kinase have consistently demonstrated significant neurological impairment in the absence of proper creatine, phosphocreatine, or creatine kinase function; thus highlighting its functional importance
chronic dosing may partially reverse neurological impairments in human CNS creatine deficiency syndromes
Several studies have suggested that creatine supplementation may also reduce oxidative DNA damage and brain glutamate levels in Huntington disease patients
Another study highlighted that creatine supplementation marginally improved indices of mood and reduced the need for increased dopaminergic therapy in patients with Parkinson's disease
Tynor Functional Knee Support
Functional knee Support is an anterior opening device, which offers the advantage of controlled compression around the knee and a rigid lateral support and immobilization. It allows normal flexion and free movement of the knee joint.
Anterio Open able
Easy application
Controlled compression.
Perfect lateral splinting.
Anatomical design.
Tynor Functional Knee Support Features
Bi axial heavy duty aluminum hinge
Mimics the natural knee joint
Ensures full weight bearing.
Allows free flexion movement
Four way stretchable fabric
Controlled and comfortable compression
No buckling
No vaso constriction
Enhanced comfort
Open patella design
Release patellar pressure
Hold the patella in position
Can be used for Patellofemoral diseases
Wrap design with anterior closing
Easy application and removal on swollen or asymmetric knees
Easy application and removal for weak or geriatric patients.
Allows customized compression
Offers flexibility in sizing
Ergonomic design
Anti tourniquet effect - ensures no constriction to blood flow
Better grip of the product to the body.
Anatomic construction- Better functionality and Snug fit.
GCs induce increased cellular expression of receptors for several pro-inflammatory cytokines including interleukin (IL)-1 (Spriggs et al. 1990), IL-2 (Wiegers et al. 1995), IL-4 (Paterson et al. 1994), IL-6 (Snyers et al. 1990), and IFN-g (Strickland et al. 1986), as well as GM-CSF
GCs have also been shown to stimulate effector cell functions including phagocytosis by monocytes (van der Goes et al. 2000), effector cell proliferative responses (Spriggs et al. 1990), macrophage activation (Sorrells and Sapolsky 2010), and a delay of neutrophil apoptosis
a concentration- and time-dependent range of GC effects that are both pro- and anti-inflammatory
basal (diurnal) concentrations of cortisol do not exert an anti-inflammatory effect on several pro-and anti-inflammatory mediators of the human immune inflammatory response
withdrawal of cortisol activity in vivo did not lead to increased inflammatory responsiveness of immune effector cells
maximal suppression of inflammation was achieved by a stress-associated, but still physiologic, cortisol concentration. There was no greater anti-inflammatory effect at higher cortisol concentrations (Yeager et al. 2005) although IL-10 concentrations continued to increase with increasing cortisol concentrations as we and others have shown
acutely, physiological cortisol concentrations are anti-inflammatory and, as proposed, act to limit over expression of an inflammatory response that could lead to tissue damage
Acutely, cortisol has anti-inflammatory effects following a systemic inflammatory stimulus (Figure 4). However, a cortisol concentration that acts acutely to suppress systemic inflammation also has a delayed effect of augmenting the inflammatory response to subsequent, delayed stimulu
1) GCs can exert pro-inflammatory effects on key inflammatory processes and, 2) GC regulation of inflammation can vary from anti- to a pro-inflammatory in a time-dependent manner
The immediate in vivo effect of both stress-induced and pharmacological GC concentrations is to suppress concurrent inflammation and protect the organism from an excessive or prolonged inflammatory response
GCs alone, in the absence of an inflammatory stimulus, up-regulate monocyte mRNA and/or receptors for several molecules that participate in pro-inflammatory signaling, as noted above and in the studies presented here.
In humans, as shown here, if in vivo GC concentrations are elevated concurrent with an inflammatory stimulus, anti-inflammatory effects are observed
In sharp contrast, with a time delay of 12 or more hours between an increased GC concentration and the onset of an inflammatory stimulus, enhancing effects on inflammation are observed. These effects have been shown to persist in humans for up to 6 days
GC-induced enhancement of inflammatory responses is maximal at an intermediate concentration, in our studies at a concentration that approximates that observed in vivo following a major systemic inflammatory stimulus
In addition to enhanced responses to LPS, recently identified pro-inflammatory effects of GCs also show enhanced localization of effector cells at inflammatory sites
we hypothesize that pre-exposure to stress-associated cortisol concentrations “prime” effector cells of the monocyte/macrophage lineage for an augmented pro-inflammatory response by; a) inducing preparative changes in key regulators of LPS signal transduction, and b) enhancing localization of inflammatory effector cells at potential sites of injury
IR also indirectly induces DNA damage by stimulating reactive oxygen species (ROS) production
IR is known to induce EMT in vitro
p53 is activated in response to IR-induced DNA damage
IR paradoxically also promotes tumour recurrence and metastasis
DNA double-strand breaks (DSBs)
cancer cells undergoing EMT acquire invasive and metastatic properties
changes in the tumour microenvironment (TME)
IR seems to induce EMT and CSC phenotypes by regulating cellular metabolism
EMT, stemness, and oncogenic metabolism are known to be associated with resistance to radiotherapy and chemotherapy
Hanahan and Weinberg proposed ten hallmarks of cancer that alter cell physiology to enhance malignant growth: 1) sustained proliferation, 2) evasion of growth suppression, 3) cell death resistance, 4) replicative immortality, 5) evasion of immune destruction, 6) tumour-promoting inflammation, 7) activation of invasion and metastasis, 8) induction of angiogenesis, 9) genome instability, and 10) alteration of metabolism
EMT is a developmental process that plays critical roles in embryogenesis, wound healing, and organ fibrosis
IR is known to induce stemness and metabolic alterations in cancer cells
transforming growth factor-β [TGF-β], epidermal growth factor [EGF]) and their associated signalling proteins (Wnt, Notch, Hedgehog, nuclear-factor kappa B [NF-κB], extracellular signal-regulated kinase [ERK], and phosphatidylinositol 3-kinase [PI3K]/Akt
activate EMT-inducing transcription factors, including Snail/Slug, ZEB1/δEF1, ZEB2/SIP1, Twist1/2, and E12/E47
Loss of E-cadherin is considered a hallmark of EMT
IR has been shown to induce EMT to enhance the motility and invasiveness of several cancer cells, including those of breast, lung, and liver cancer, and glioma cells
IR may increase metastasis in both the primary tumour site and in normal tissues under some circumstance
sublethal doses of IR have been shown to enhance the migratory and invasive behaviours of glioma cells
ROS are known to play an important role in IR-induced EMT
High levels of ROS trigger cell death by causing irreversible damage to cellular components such as proteins, nucleic acids, and lipids, whereas low levels of ROS have been shown to promote tumour progression—including tumour growth, invasion, and metastasis
hypoxia-inducible factor-1 (HIF-1) is involved in IR-induced EMT
Treatment with the N-acetylcysteine (NAC), a general ROS scavenger, prevents IR-induced EMT, adhesive affinity, and invasion of breast cancer cells
Snail has been shown to play a crucial role in IR-induced EMT, migration, and invasion
IR activates the p38 MAPK pathway, which contributes to the induction of Snail expression to promote EMT and invasion
NF-κB signalling that promotes cell migration
ROS promote EMT to allow cancer cells to avoid hostile environments
HIF-1 is a heterodimer composed of an oxygen-sensitive α subunit and a constitutively expressed β subunit.
Under normoxia, HIF-1α is rapidly degraded, whereas hypoxia induces stabilisation and accumulation of HIF-1α
levels of HIF-1α mRNA are enhanced by activation of the PI3K/Akt/mammalian target of rapamycin (mTOR)
IR is known to increase stabilisation and nuclear accumulation of HIF-1α, since hypoxia is a major condition for HIF-1 activation
IR induces vascular damage that causes hypoxia
ROS is implicated in IR-induced HIF-1 activation
IR causes the reoxygenation of hypoxic cancer cells to increase ROS production, which leads to the stabilisation and nuclear accumulation of HIF-1
IR increases glucose availability under reoxygenated conditions that promote HIF-1α translation by activating the Akt/mTOR pathway
The stabilised HIF-1α then translocates to the nucleus, dimerizes with HIF-1β, and increases gene expression— including the expression of essential EMT regulators such as Snail—to induce EMT, migration, and invasion
TGF-β signalling has been shown to play a crucial role in IR-induced EMT
AP-1 transcription factor is involved in IR-induced TGF-β1 expression
Wnt/β-catenin signalling is also implicated in IR-induced EMT
Notch signalling is known to be involved in IR-induced EMT
IR also increases Notch-1 expression [99]. Notch-1 is known to induce EMT by upregulating Snail
PAI-1 signalling is also implicated in IR-induced Akt activation that increases Snail levels to induce EMT
EGFR activation is known to be associated with IR-induced EMT, cell migration, and invasion by activating two downstream pathways: PI3K/Akt and Raf/MEK/ERK
ROS and RNS are also implicated in IR-induced EGFR activation
IR has also been shown to activate Hedgehog (Hh) signalling to induce EMT
IR has been shown to induce Akt activation through several signalling pathways (EGFR, C-X-C chemokine receptor type 4 [CXCR4]/C-X-C motif chemokine 12 [CXCL12], plasminogen activator inhibitor 1 [PAI-1]) and upstream regulators (Bmi1, PTEN) that promote EMT and invasion
CSCs possess a capacity for self-renewal, and they can persistently proliferate to initiate tumours upon serial transplantation, thus enabling them to maintain the whole tumour
Conventional cancer treatments kill most cancer cells, but CSCs survive due to their resistance to therapy, eventually leading to tumour relapse and metastasis
identification of CSCs, three types of markers are utilised: cell surface molecules, transcription factors, and signalling pathway molecules
CSCs express distinct and specific surface markers; commonly used ones are CD24, CD34, CD38, CD44, CD90, CD133, and ALDH
Transcription factors, including Oct4, Sox2, Nanog, c-Myc, and Klf4,
signalling pathways, including those of TGF-β, Wnt, Hedgehog, Notch, platelet-derived growth factor receptor (PDGFR), and JAK/STAT
microRNAs (miRNAs), including let-7, miR-22, miR-34a, miR-128, the miR-200 family, and miR-451
Non-CSCs can be reprogrammed to become CSCs by epigenetic and genetic changes
EMT-inducing transcription factors, such as Snail, ZEB1, and Twist1, are known to confer CSC properties
Signalling pathways involved in EMT, including those of TGF-β, Wnt, and Notch, have been shown to play important roles in inducing the CSC phenotype
TGF-β1 not only increases EMT markers (Slug, Twist1, β-catenin, N-cadherin), but also upregulates CSC markers (Oct4, Sox2, Nanog, Klf4) in breast and lung cancer cells
some CSC subpopulations arise independently of EMT
IR has been shown to induce the CSC phenotype in many cancers, including breast, lung, and prostate cancers, as well as melanoma
Genotoxic stress due to IR or chemotherapy promotes a CSC-like phenotype by increasing ROS production
IR has been shown to induce reprogramming of differentiated cancer cells into CSCs
In prostate cancer patients, radiotherapy increases the CD44+ cell population that exhibit CSC properties
IR also induces the re-expression of stem cell regulators, such as Sox2, Oct4, Nanog, and Klf4, to promote stemness in cancer cells
EMT-inducing transcription factors and signalling pathways, including Snail, STAT3, Notch signalling, the PI3K/Akt pathway, and the MAPK cascade, have been shown to play important roles in IR-induced CSC properties
STAT3 directly binds to the Snail promoter and increases Snail transcription, which induces the EMT and CSC phenotypes, in cisplatin-selected resistant cells
Other oncogenic metabolic pathways, including glutamine metabolism, the pentose phosphate pathway (PPP), and synthesis of fatty acids and cholesterol, are also enhanced in many cancers
metabolic reprogramming
HIF-1α, p53, and c-Myc, are known to contribute to oncogenic metabolism
metabolic reprogramming
tumour cells exhibit high mitochondrial metabolism as well as aerobic glycolysis
occurring within the same tumour
CSCs can be highly glycolytic-dependent or oxidative phosphorylation (OXPHOS)-dependen
mitochondrial function is crucial for maintaining CSC functionality
cancer cells depend on mitochondrial metabolism and increase mitochondrial production of ROS that cause pseudo-hypoxia
HIF-1 then enhances glycolysis
CAFs have defective mitochondria that lead to the cells exhibiting the Warburg effect; the cells take up glucose, and then secrete lactate to 'feed' adjacent cancer cells
Epithelial cancer cells express MCT1, while CAFs express MCT4. MCT4-positive, hypoxic CAFs secrete lactate by aerobic glycolysis, and MCT1-expressing epithelial cancer cells then uptake and use that lactate as a substrate for the tricarboxylic acid (TCA) cycle
MCT4-positive cancer cells depend on glycolysis and then efflux lactate, while MCT1-positive cells uptake lactate and rely on OXPHOS
metabolic heterogeneity induces a lactate shuttle between hypoxic/glycolytic cells and oxidative/aerobic tumour cells
bulk tumour cells exhibit a glycolytic phenotype, with increased conversion of glucose to lactate (and enhanced lactate efflux through MCT4), CSC subsets depend on oxidative phosphorylation; most of the glucose entering the cells is converted to pyruvate to fuel the TCA cycle and the electron transport chain (ETC), thereby increasing mitochondrial ROS production
the major fraction of glucose is directed into the pentose phosphate pathway, to produce redox power through the generation of NADPH and ROS scavengers
HIF-1α, p53, and c-Myc, are known to contribute to oncogenic metabolism
regulatory molecules involved in EMT and CSCs, including Snail, Dlx-2, HIF-1, STAT3, TGF-β, Wnt, and Akt, are implicated in the metabolic reprogramming of cancer cells
HIF-1 induces the expression of glycolytic enzymes, including the glucose transporter GLUT, hexokinase, lactate dehydrogenase (LDH), and MCT, resulting in the glycolytic switch
HIF-1 represses the expression of pyruvate dehydrogenase kinase (PDK), which inhibits pyruvate dehydrogenase (PDH), thereby inhibiting mitochondrial activity
STAT3 has been implicated in EMT-induced metabolic changes as well
TGF-β and Wnt play important roles in the metabolic alteration of cancer cells
Akt is also implicated in the glycolytic switch and in promoting cancer cell invasiveness
EMT, invasion, metastasis, and stemness
pyruvate kinase M2 (PKM2), LDH, and pyruvate carboxylase (PC), are implicated in the induction of the EMT and CSC phenotypes
decreased activity of PKM2 is known to promote an overall shift in metabolism to aerobic glycolysis
LDH catalyses the bidirectional conversion of lactate to pyruvate
High levels of LDHA are positively correlated with the expression of EMT and CSC markers
IR has been shown to induce metabolic changes in cancer cells
IR enhances glycolysis by upregulating GAPDH (a glycolysis enzyme), and it increases lactate production by activating LDHA, which converts pyruvate to lactate
IR enhances glycolysis by upregulating GAPDH (a glycolysis enzyme), and it increases lactate production by activating LDHA, which converts pyruvate to lactate
IR also elevates MCT1 expression that exports lactate into the extracellular environment, leading to acidification of the tumour microenvironment
IR increases intracellular glucose, glucose 6-phosphate, fructose, and products of pyruvate (lactate and alanine), suggesting a role for IR in the upregulation of cytosolic aerobic glycolysis
Lactate can activate latent TGF-
lactate stimulates cell migration and enhances secretion of hyaluronan from CAF that promote tumour metastasis
promote tumour survival, growth, invasion, and metastasis; enhance the stiffness of the ECM; contribute to angiogenesis; and induce inflammation by releasing several growth factors and cytokines (TGF-β, VEGF, hepatocyte growth factor [HGF], PDGF, and stromal cell-derived factor 1 [SDF1]), as well as MMP
tumours recruit the host tissue’s blood vessel network to perform four mechanisms: angiogenesis (formation of new vessels), vasculogenesis (de novo formation of blood vessels from endothelial precursor cells), co-option, and modification of existing vessels within tissues.
immunosuppressive cells such as tumour-associated macrophages (TAM), MDSCs, and regulatory T cells, and the immunosuppressive cytokines, TGF-β and interleukin-10 (IL-10)
immunosuppressive cells such as tumour-associated macrophages (TAM), MDSCs, and regulatory T cells, and the immunosuppressive cytokines, TGF-β and interleukin-10 (IL-10)
intrinsic immunogenicity or induce tolerance
cancer immunoediting’
three phases: 1) elimination, 2) equilibrium, and 3) escape.
The third phase, tumour escape, is mediated by antigen loss, immunosuppressive cells (TAM, MDSCs, and regulatory T cells), and immunosuppressive cytokines (TGF-β and IL-10).
IR can elicit various changes in the TME, such as CAF activity-mediated ECM remodelling and fibrosis, cycling hypoxia, and an inflammatory response
IR activates CAFs to promote the release of growth factors and ECM modulators, including TGF-β and MMP
TGF-β directly influences tumour cells and CAFs, promotes tumour immune escape, and activates HIF-1 signalling
IR also promotes MMP-2/9 activation in cancer cells to promote EMT, invasion, and metastasis
IR-induced Snail increases MMP-2 expression to promote EMT
Radiotherapy has the paradoxical side-effect of increasing tumour aggressiveness
IR promotes ROS production in cancer cells, which may induce the activation of oncogenes and the inactivation of tumour suppressors, which further promote oncogenic metabolism
Metabolic alterations
oncogenic metabolism
elicit various changes in the TME
Although IR activates an antitumour immune response, this signalling is frequently suppressed by tumour escape mechanisms
subnormal plasma ascorbic acid levels are a predictable feature in patients with severe sepsis
Ascorbic acid depletion in sepsis results from ascorbic acid consumption by the reduction of plasma free iron, ascorbic acid consumption by the scavenging of aqueous free radicals (peroxyl radicals), and by the destruction of the oxidized form of ascorbic acid dehydroascorbic acid
Sepsis further inhibits intracellular reduction of dehydroascorbic acid, producing acute intracellular ascorbic acid depletion
Ascorbic acid treated patients in this study exhibited rapid and sustained increases in plasma ascorbic acid levels using an intermittent every six hours administration protocol
Septic ascorbic acid-deficient neutrophils fail to undergo normal apoptosis. Rather, they undergo necrosis thereby releasing hydrolytic enzymes in tissue beds, thus contributing to organ injury
We speculate that intravenous ascorbic acid acts to restore neutrophil ascorbic acid levels
Repletion of ascorbic acid in this way allows for normal apoptosis, thus, preventing the release of organ damaging hydrolytic enzymes.
Study finds IV vitamin C in patients with sepsis is very safe and blunts the effects (endothelial damage, end organ damage...) of sepsis. Of note, the IV vitamin C group reached serum levels of ascorbic acid of 1,592 to 5,722 micromol/L. The IV groups maintained elevated serum C levels for up to 96 hours post infusion.
daily doses up to 400 mg of HCQ or 250 mg CQ for several years are considered to carry an acceptable risk for CQ-induced retinopathies, with the exception of individuals of short stature
chronic CQ or HCQ therapy be monitored through regular ophthalmic examinations (3–6 month intervals), full blood counts and blood glucose level checks
long-term HCQ exposure, skeletal muscle function and tendon reflexes should be monitored for weakness
both CQ and HCQ, specific caution is advised in patients suffering from impaired hepatic function (especially when associated with cirrhosis), porphyria, renal disease, epilepsy, psoriasis, glucose-6-phosphate dehydrogenase deficiency and known hypersensitivity to 4-aminoquinoline compounds
CQ and HCQ can effectively increase the efficacy of various anti-cancer drugs
CQ can prevent the entrapment of protonated chemotherapeutic drugs by buffering the extracellular tumour environment and intracellular acidic spaces
This study recommends an adjuvant HCQ dose of 600 mg, twice daily.
HCQ addition was shown to produce metabolic stress in the tumours
HCQ (400 mg/day)
important effects of CQ and HCQ on the tumour microenvironment
The main and most studied anti-cancer effect of CQ and HCQ is the inhibition of autophagy
the expression levels of TLR9 are higher in hepatocellular carcinoma, oesophageal, lung, breast, gastric and prostate cancer cells as compared with adjacent noncancerous cells, and high expression is often linked with poor prognosis
TLR9-mediated activation of the NF-κB signalling pathway and the associated enhanced expression of matrix metalloproteinase-2 (MMP-2), MMP-7 and cyclo-oxygenase 2 mRNA
HCQ can activate caspase-3 and modulate the Bcl-2/Bax ratio inducing apoptosis in CLL, B-cell CLL and glioblastoma cells
In triple-negative breast cancer, CQ was shown to eliminate cancer stem cells through reduction of the expression of Janus-activated kinase 2 and DNA methyl transferase 1 [106] or through induction of mitochondrial dysfunction, subsequently causing oxidative DNA damage and impaired repair of double-stranded DNA breaks
CQ or HCQ would be considered for use in combination with immunomodulation anti-cancer therapies
Therapies used in combination with CQ or HCQ include chemotherapeutic drugs, tyrosine kinase inhibitors, various monoclonal antibodies, hormone therapies and radiotherapy
Most studies hypothesise that CQ and HCQ could increase the efficacy of other anti-cancer drugs by blocking pro-survival autophagy.
daily doses between 400 and 1200 mg for HCQ are safe and well tolerated, but two studies identified 600-mg HCQ daily as the MTD
HCQ is often administered twice daily to limit plasma fluctuations and toxicity

 Nathan Goodyear on 23 May 16study finds severe hypothyroidism that is abrupt is not associated with low sodium.
Nathan Goodyear on 23 May 16study finds severe hypothyroidism that is abrupt is not associated with low sodium. Group items tagged
Group items tagged